Появление, расцвет и исчезновение мамонтов связывают прочной нитью начало кайнозойской эры с нашими днями в длинную, интересную, сложную и не всегда ясную историю.
Наше представление о ней изменяется при новых находках. А находки – это всегда новые загадки. Путь хоботных во времени и пространстве стал повествовательной нитью рассказа Naked Science об этих животных.

История мамонтов началась задолго до их появления — с возникновением первых хоботных. Отряд хоботных (Proboscidea) существует многие десятки миллионов лет, породив множество групп, заселявших просторы разных континентов. Эта демонстрация эволюционной успешности росла с приближением к нашему времени. Однако эволюционная удача не вечна. Мамонты, один из венцов эволюции хоботных, ярко расцвели во время последнего оледенения. И даже пережили его окончание, угаснув совсем недавно, уже в историческое время человека. Мамонты оставили нам не только хорошо сохранившиеся в вечной мерзлоте туши, но и захватывающую интригу своего появления и исчезновения.
Определить точное время появления на Земле хоботных невозможно, как и указать день, до которого они не были хоботными, а после – вдруг уже таковыми стали. Плавные непрерывные изменения форм от одной к другой не могут иметь четких, абсолютных границ в потоке времени. К тому же мы опираемся на находки, тогда как реальные события разделения форм животных всегда древнее попавшего в руки образца результата разделения. И эти находки тоже относительны: их датировки не абсолютно точны, а оцениваемые размеры и вес существ сильно различаются. Отнесение к группам, выделение видов и родов постоянно настраивается и пересматривается с новыми находками и методами, меняя классификацию и взгляды на родственные связи. Отдавая себе в этом отчет и понимая некоторую относительность всех оценок, рассмотрим эволюцию баллистики хоботных и траектории мамонтов в ней.
Зажигание, первая ступень — мелкие
Первое обнаружение движения хоботных произошло на глубокой временной шкале. 60 млн. Много лет назад, в палеоцене первого геологического периода кайнозойской эры, жили первые хоботные. В первой эпохе этой эры, которая длилась с 66 по 56 миллионов лет назад, известен район расположения боевой стартовой позиции БСП: это сегодняшний фосфатный бассейн Оулед Абдун в середине Марокко. Оттуда происходят фрагменты челюстей двух небольших животных. Первым из обнаруженных был эритерий размером с маленькую собаку.
Второе животное, названное фосфатерием, обитало в районе старта, где обнаружены фосфатные шахты. 58 млн. Несколько миллионов лет назад, после первого вида, появился второй. Он был чуть больше предшественника, но в три раза тяжелее и по размерам напоминал крупную собаку. У обоих ещё не было хоботов, но резцы, будущие бивни, уже заметно увеличились. Вышедшие из мелких насекомоядных, эти два вида, вероятно, всё еще питались разнообразной пищей, пока не специализировались на растительной. Поэтому зубы у них были мелкими с небольшими бугорками эмалевых коронок, предназначенными для разгрызания мелких насекомых.
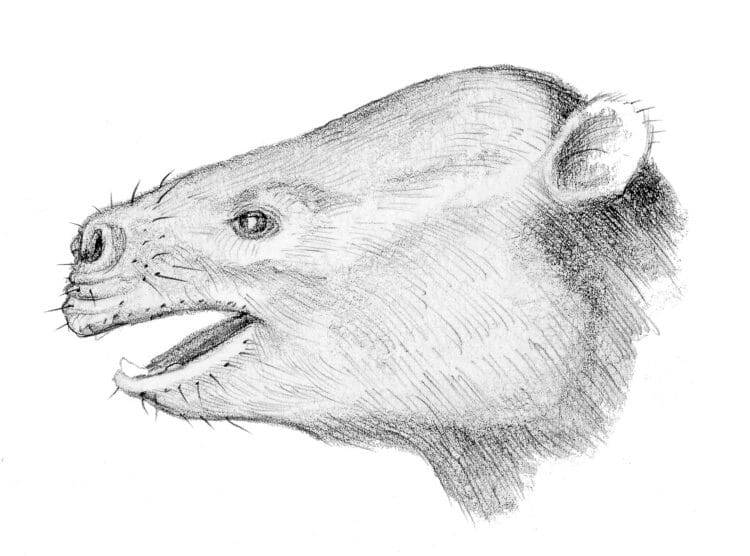
Через десяток миллионов лет после… . 45 млн. Несколько миллионов лет назад, в среднем эоцене (вторая эпоха палеогена, 56-33,9 млн. лет назад), по северу Африки широко распространились родственники первых двух, объединяясь в обширную группу нумидотериев (Numidotherium).
Те территории, где жили нумидотерии, простирались от современного Марокко до Алжира, Ливии, Туниса, позже ставших землями древнего Нумидийского царства.
Размеры нумидотериев варьировались от габаритов средней собаки до большой грузной свиньи, крупнейшие вырастали и со среднюю корову. Благодаря полуводному образу жизни у них отрастил небольшой мускульный хобот из носа и верхней губы, подобно современным тапирам.
Нумидотерии исчезли примерно 30 млн. лет назад, в раннем олигоцене.
Примерно в одно время, но чуть позже, жили схожие по размеру и строению меритерии. С расцветом на отметках 37-35 млн. Несколько миллионов лет назад его выделили в отдельное семейство. Назвали по месту жительства – у озера Меридова в Египте. С длинным, низким телом и полуводным образом жизни меритерий продвинулся южнее, до современных Мали и Сенегала. Увеличенные резцы предсказывали будущие большие бивни хоботных. По зубам видно, что питались они мягкой водной растительностью (которую меритерий собирал коротким, как у тапира, хоботом). Меритерии просуществовали до середины олигоцена (последняя эпоха палеогена, 33,9 — 23 млн лет назад), исчезнув около 30-28 млн лет назад.

Первая ступень хоботных, ступень ранних мелких форм, отработала и отделилась. Запустился 60Мільйони років тому рання лосхотних пройшли половину загального еволюційного шляху. 30Несколько миллионов лет назад ранние хоботные постоянно увеличивались в размерах в ходе эволюции.
Прирост массы тела осуществлялся за счет использования широко доступных топлив — пищевых ресурсов. Первоначальная всеядность постепенно замещалась на самый распространенный и богатый тип пищи — растительность. Для упрощения поедания (подобно сжатию поступающего потока воздухом вбирающем устройства сверхзвукового самолета) мягкие водные растения собирались в пучок.
Возня при таком сборе формировала мягкий орган захвата — короткий хобот, закрепляя переход на растительную пищу. Многочисленные зубы сдавливали мягкую массу водных растений своими универсальными небольшими бугорками эмалевых коронок, пока еще не специализированными под крупную или твердую растительную пищу. Увеличение отдельных резцов и растущая увесистость тел были показателями перехода к последующим большим формам с хоботом, бивнями и большими размерами.
Вторая ступень — крупные
Эоцен (56-33,9 млн лет назад), вторая эпоха первого геологического периода палеогена кайнозойской эры, продолжался 22 миллиона лет. В начале Эпохи климат был самым теплым в истории кайнозоя. Позже установилось долгое и усиливающееся похолодание, которое не смягчались короткими потеплениями. К концу Эоцена охлаждение ускорилось, а ледниковый щит Антарктиды быстро рос. Быстрые изменения климата привели к заметному вымиранию растений и животных. Это событие, хоть и слабее пяти великих массовых вымираний, было настолько значительным, что стало границей между второй и третьей эпохами палеогена — эоцен завершился, начался олигоцен. Изменения растительного и животного мира в Европе были столь серьезны, что получили название Великого перелома.
Однако хоботные преодолели этот климатический рубеж (33,9 млн лет назад) без проблем и только укрепились.
Их эволюционная гибкость и маневренность победили и выросли. В результате возникли хоботные нового типа — большие, массивная животные с хоботом, бивнями и новыми преимуществами для эволюции. Так началась следующая часть пути хоботны, полная событий и обильная формами.
Вторая ступень эволюции разгоняла хоботных. На участке второй ступени шла активная работа. Эволюция проверила пластичность хоботных на отклонениях от основной траектории, поработав с материалом в разных местностях и метеоусловиях. Большая по массе, универсальная живая фабрика переработки растительности легко прокладывала себе путь, торя эволюционную дорогу. А та привела к стремительно развивающимся крупнотоннажным семействам, из которых стоит отметить следующую пятерку.
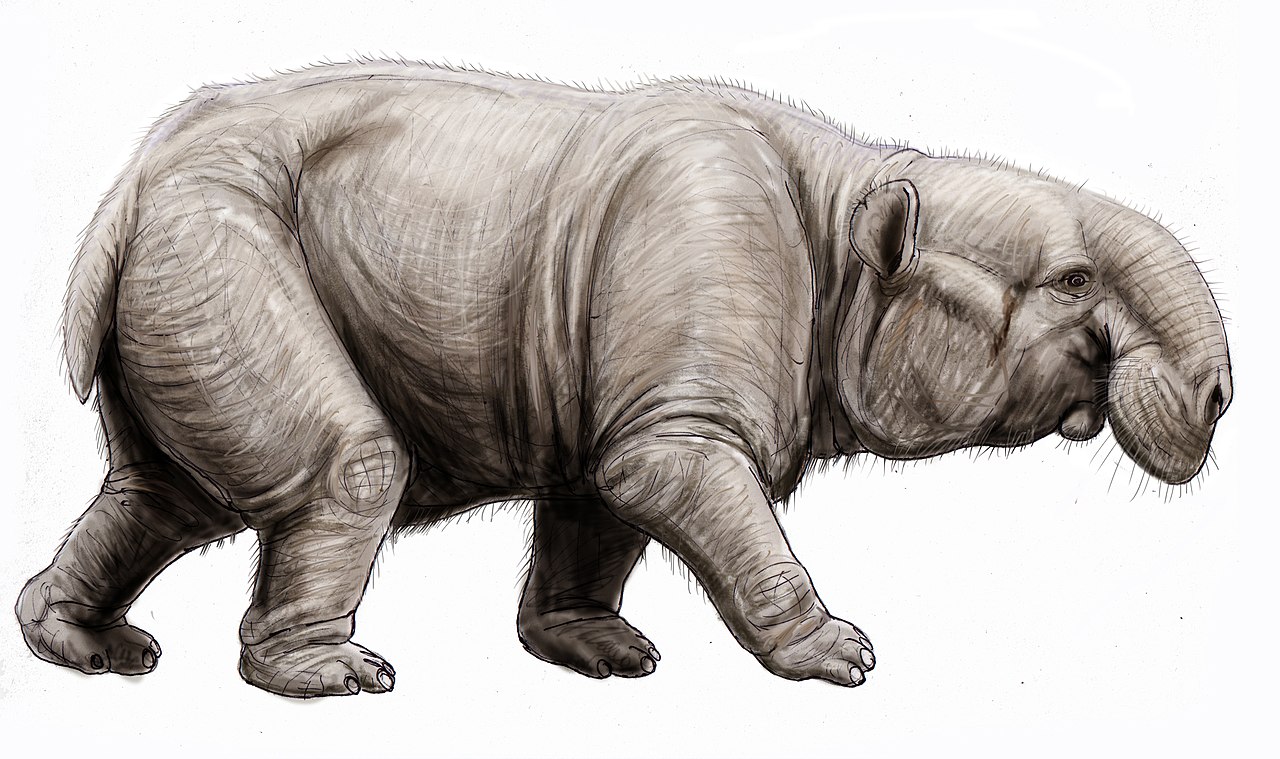
35-30 млн. Несколько миллионов лет назад, с позднего эоцена по ранний олигоцен, в северной Африке появились и рассеялись первые крупные и тяжелые хоботные, что отразилось в их названии. Семейство баритериев (Barytheriidae) получило такое название от греческого «барис» — «тяжесть». Это слово напоминает баллистическое понятие барицентра Земли — центра, вокруг которого обращаются по орбитам спутники. Тяжелый зверь достигал размеров и массы некрупных современных слонов. В верхней и нижней челюсти баритерий нес по паре уже довольно крупных резцов, предшественников бивней. Хобот его был пока небольшой. Главное новое качество — крупные габариты корпуса и большая масса. Это уменьшило давление хищников, облегчив дальнейшую эволюцию.
В это же время, около 35 млн. Летом назад появился палеомастодонт (Palaeomastodon). Представители его семейства были ещё крупнее и массивнее, превышали пару метров в холке, вес «изделия» приближался к трем тоннам. Жители северной Африки. Хобот палеомастодонтов оставался ещё небольшим, но уже сильно укрупнились верхние и нижние резцы, разделившись по форме. В удлиненной нижней челюсти они стали плоскими, как две лопатки или вытянутых ложки. Вероятно, служили для выкапывания растений из полузатопленных болотистых грунтов. Такая практика переместила уши ближе к макушке, во избежание заливания их водой при неглубоких погружениях головы. Пролетев по траектории свои 10 млн лет, семейство палеомастодонтов исчезло 25 млн. лет назад.

В середине их пути, 30Миллионы лет назад от палеомастодонов произошли предки семейства гребнезубых мастодонтов, или просто мастодонтов (Mammutidae). Крупные хоботные с четырьмя большими бивнями позже утратили бивни нижней челюсти. Название получили от сестринского семейства бугорчатозубых, от которого были отделены, унаследовав общее название. Высокие и островерхие жевательные гребни поперек зубов располагались несколькими рядами, образуя гребнезубую дробилку для крупного материала. Гребни на зубе напоминали лезвия ломов или колунов, с чуть выраженной впадиной посередине гребня. Эта машина дробления означает переработку крупных веток и лес как среду обитания.
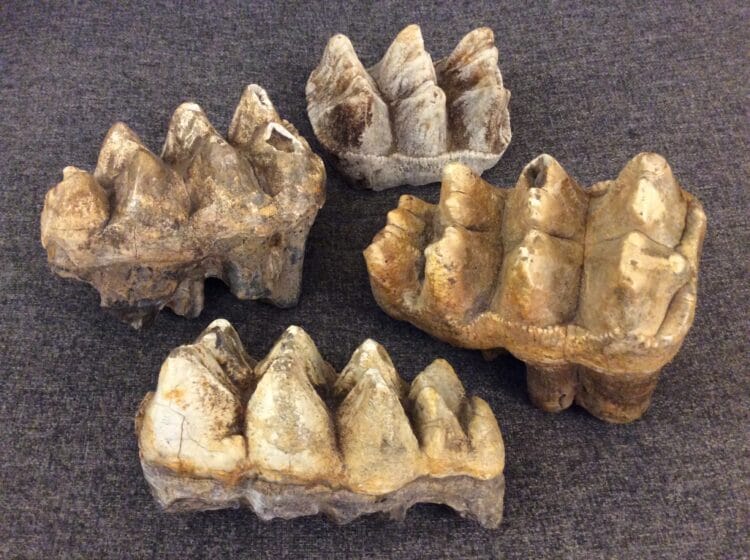
Гребензубые мастодонты сформировали множество видов, около двадцати из которых известно сегодня.
Выглядели животные приземистыми вытянутыми хоботными с длинными прямыми бивнями. Удлиненное и относительно невысокое тело облегчало им путь сквозь скопления крупных деревьев.
Эти лесные танки расселились по трем континентам: Африке, Евразии и Северной Америке. Последние мастодонты вымерли в Северной Америке с завершением ледникового периода около 10 тысяч лет назад, в том числе под ударами копий палеоамериканского населения. Расселились звери не только по северным зонам континента, но и в субтропические леса юга.
Мастодонты хорошо изучены. В американской мерзлоте найдены замерзшие туши и их части, подобно мамонтовым. Обитали последние из мастодонтовых могикан в лесах и болотисто-лесной местности. Известны черепа мастодонтов со следами ударов палеоиндейским копьем. Daniel FisherВедущий специалист планеты по вопросам вымирания американских мастодонтов, а также мамонтов Евразии и Америки собрал большой объем уникальных данных об их исчезновении. Продолжаются уточнения роли в этом вымирании человека и климата.

25 миллионов Много лет назад, в позднем олигоцене, от общего пучка траекторий эволюции отделилось боковое ответвление динотериев, или дейнотериев (Deinotherium). «Ужасные звери» (так их название переводится с древнегреческого) со временем расселились по Европе, Средней Азии и северу Индостана. Три рода динотериев сменяли друг друга в хронологическом порядке. Начавшись с небольшого полуторатонного формата, около 1,5 тонн и 2 метров высоты, эти хоботные укрупнились до позднего динотерия гигантского. Он превосходил ростом всех современных ему хоботных, и только в конце своего существования уступил самым крупным мамонтам-исполинам. Динотерий гигантский считается третьим по размерам среди всех наземных млекопитающих за всю их историю, сразу после рекордных исполинов – крупнейших слонов и индрикотериев.

У динотерия в челюсти было несколько зубов с каждой стороны, поэтому зубы были короткими. Каждый зуб имел два больших поперечных гребня, из которых один делился на два больших бугра по сторонам зуба. Огромный зуб превосходил по ширине все остальные зубные формы хоботных этого времени. Это была, вероятно, дробилка с самыми крупными зубными выступами у хоботных, и свидетельствовала о крупнодревесном характере корма — гигантский динотерий откусывал от больших деревьев очень большие куски. Самым характерным отличием динотерия была внешность: два больших бивня круто выгибались из нижней челюсти к земле и назад, к передним ногам. Как их использовал динотерий, неизвестно; возможно, обдирал кору с деревьев. Динотерии исчезли 2 млн. лет назад в начале оледенения, не пережив усиливающихся холодов.

С исчезновением гребнезубых мастодонтов и палеомастодонтов на смену им пришли гомфотерии, которые стали преобладающим видом хоботных. 15 миллионов Лет назад существовала обширная группа с разнообразными предками и не всегда ясными эволюционными путями, включавшая несколько семейств и родов, иногда с причудливыми формами — амебелодоны и платибелодоны, трилофодоны и тетралофодоны, эубелодоны и ананкусы, также называемые овернскими мастодонтами, и другие. Некоторые гомфотерии были больше похожи на гребнезубых мастодонтов, другие напоминали слонов. Название всей группе дано роду гомфотериев, или бугорчатозубых мастодонтов.
Жевательная поверхность зуба состояла из двух рядов высоких округлых бугорков, похожих на ряды сросшихся сосков коровьего вымени — или сложенных в троеперстие пальцев. Повторяющиеся V-образные ряды напоминают тракторный протектор колеса «Кировца» или «Беларуси». По этому признаку бугорчатозубые мастодонты получили название мастодонтов, от латинского «маст» — сосок и «донт» — зуб. Позже их назвали гомфотериями за взаимное расклинивание их четырех бивней: нижние входили в зацепление с верхними, как английская булавка или античная бронзовая заколка-фибула. Гомфотерий – «зверь с заколкой (бивневой)». Около 5 миллионов лет назад их постепенно на земных равнинах сменили слоны и гребнезубые мастодонты. Но эволюционная гибкость гомфотериев была велика; они выживали множеством форм, и самые поздние виды дожили до конца ледникового периода и, как считается, даже встретились с человеком в Южной Америке.

12 миллионов Несколько миллионов лет назад, начиная с середины миоцена, от гомфотериев отделились стегодоны (Stegodontidae). Часть названия «стего» означает «покрытый», то есть зуб с крышей. Стегодоны очень напоминали своих потомков — слонов. Их длинные изогнутые бивни смотрели прямо вперед и шли практически параллельно, почти смыкаясь концами. Размеры стегодонов менялись от крупных до уменьшающихся форм на островах, куда, видимо, стегодоны заплывали, так как в те времена к этим островам не было сухопутных перешейков.
Что такое покрытый зуб и чем важна эта «крыша»?
На протяжении всей своей истории развития хоботные сохраняли одну и ту же зубную архитектуру, которая была общей для всех семейств этого отряда. Жевательная поверхность располагалась на толстой цельной коронке из эмали, самого твердого вещества позвоночных, впервые появившегося в девоне на кожных «зубах» акул — твердых плакоидных чешуях. Зубная крышка из эмали концентрирует силу жевательных мышц на поверхностных буграх и гребнях. Концентрация необходима для разрушения прочной пищи, преодоления её пределов прочности. Крышка полностью покрывала всю рабочую, жевательную сторону зуба, подобно короне голове королей. Поэтому эмалевая покрышка зуба получила название «коронка».
Под эмалью находился более мягкий, но все же прочный костный дентин. Ниже располагались корни зуба с каналами нервно-сосудистых пучков, обеспечивавших питание зубных тканей. Такое строение зубов свойственно большинству млекопитающих, в том числе и человеку. Человек воспроизвел подобную архитектуру в технике, применяя твердосплавные напайки на измельчающий и режущий инструмент — искусственные зубы сверл и резцов, фрез и буров, долот и проходческих щитов.


Вторая ступень завершила работу, разогнав эволюцию древних хоботных на второй половине пути к крупным формам и широкому распространению по различным средам обитания. В общем направлении движения во времени появилось много других семейств, а также множество родов и видов, большая часть которых остаётся не упомянутой, поскольку точная классификация их – сложный и бесконечный процесс.
Эстафеты форм хоботных прослеживаются не всегда чётко и достаточно условно, а преемственность относительна. Чаще можно говорить о том, что в определённые периоды времени виднелись характерные семейства. Для некоторых линий и отрезков времени преемственность более явная.
Ступень разведения — слоны
Слоны появились на Земле около 7 млн.Несколько родов, образовав семейство Elephantidae лет назад, расходились по своим траекториям, как боеголовки от ступени разведения межконтинентальной баллистической ракеты. С тем лишь различием, что боеголовки расходятся в начале основной траектории, с началом свободного полета в космосе; слоны же возникли, наоборот, на самой поздней части траектории хоботных. Эволюционно маневренные, слоны продвигались по любым типам ландшафтов, показывая высокую степень адаптивности. Слоны могли жить в саваннах и пустынях, в лесу и лесостепи, в степи и тундростепи. И несли в своих челюстях коренное отличие от остальных хоботных — все слоны имели один общий признак — принципиально новый тип зуба.
Эмалевые покрышки у зуба отсутствовали. Зуб слона состоял из костного зубного цемента, вытянутого вдоль челюсти. В него поперечно погружены были отдельные плоские эмалевые пластины с мягкой дентиновой сердцевиной. Пластины располагались в зубе как свинцовые в корпусе аккумуляторной батареи, длинной пачкой образуя сам зуб. Высокотвердыми торцами пластины выходили поперек жевательной поверхности зуба, образуя на ней полосатую терку. Внутри эмалевой пластины содержался более мягкий дентин, на торце пластины дававший впадинку внутреннего износа.

Бивни слонов не содержат эмали и полностью состоят из дентина, плотного костного вещества зуба, более мягкого, чем эмаль, но более эластичного. Возможно, бивни из эмали были бы слишком хрупкими, ломаясь на пути к обычной длине бивней. Твердость и прочность – это разные свойства. Именно эластичность дентина позволяла палеолитическим охотникам распрямлять вырезанные из бивня полосы в ровные копья двухметровой длины, например, те, которые найдены в погребениях палеолитической Сунгирской стоянки.
Первые зубные пачки первых слонов состояли из небольшого числа крупных, толстых и редкими эмалевых пластин. Нижние зубы имели всегда вогнутые жевательные поверхности, напоминающие лодки с рядами поперечных сидений. В процессе эволюции толщина пластин уменьшалась, их количество увеличивалось, а неровности жевательной терки зуба становились мельче, что происходило вслед за переходом животных на мелкие корма степей.

Первобытного рода слонов назвали примэлефасом, что означает «первослон». Два его известных вида напоминали гомфотериев. От первослонов отделился род с угловатым рисунком на жевательной поверхности зубов. По нему род получил имя локсодонтов — «ромбовидных зубов» — и дожил до наших дней двумя видами африканских слонов: крупным саванным и меньшим лесным.

Многочисленные острова и архипелаги служили «фабриками карликовости». Периодически снижающийся уровень моря давал сухопутный путь слонам из Европы на острова, а последующие подъемы запирали их. В результате популяции уменьшались в размерах, запуская механизмы классического островного измельчания. Такой цикл повторялся многократно, длительной чередой.
В итоге появились такие карликовые формы, как кипрский карликовый слон Palaeoloxodon cypriotes высотой 1,4 м и весом всего 200 кг, сицилийский карликовый слон Palaeoloxodon falconeri, достигавший в холке 90 см, и другие карликовые палеолоксодонты.
В открытых степях, где ландшафт неизменен, нет морских циклов, а развитие идет по прямой линии, росли исполины. Палеолоксодонт намадийский (Palaeoloxodon namadicus) стал самым крупным наземным млекопитающим в истории, превзойдя индрикотерия из олигоцена по высоте холки (до 5,5 м) и массе (до 25 тонн).

Остальные слоны, которые не были локсодонтами и палеолоксодонтами, имели зубные пластины с ровными параллельными сторонами, без ромбовидных утолщений на жевательной поверхности зуба. Род Elephas (настоящие слоны) сохранился до наших дней видом азиатского слона в его шести подвидах, начиная с индийского слона. Второй род слонов составили мамонты (Mammuthus), с высоким затылком и штопорообразным искривлением бивней. Мамонты формировались в плиоцене, около… 5 миллионов Несколько лет назад в восточной и южной Африке происходило эволюционное взаиморасположение, хронология и родственные связи мамонтов. Названия мамонтов также уточняются и иногда пересматриваются. Переходя к мамонтам, вспомним об объективной неабсолютности деталей рассказа.
Сначала появились мамонты, представляющие пару древних африканских видов. Первыми были представители вида Mammuthus subplanifrons с шестью эмалевыми пластинами в зубе, имеющими толстые округлые концы. 3 милионов Много лет назад мамонты мигрировали из Африки в Европу. Первым европейцем стал румынский мамонт, который расселился от Англии до Китая и имел во рту 8 — 10 эмалевых пластин. Благодаря широте распространения мамонты способствовали ускорению эволюции. Около 2 миллионовНесколько лет назад первого европейца вытеснил мамонт меридионалис (Mammuthus meridionalis), или южный слон, также называемый архидискодоном, с 15—17 зубными пластинами, которые становились тоньше по мере увеличения их числа в зубе.
Через полмиллиона лет его сменил могучий степной мамонт (Mammuthus trogontherii) с двадцатью или двадцатью двумя зубными пластинами, обитавший в восточноазиатских степях. миллион лет Перейдя на Аляску сушей, мамонт заселил своими потомками всю Северную Америку. В различных климатических зонах континента появились несколько видов североамериканских мамонтов: от гигантского императорского (Mammuthus imperator) до карликового островного эксилиса (Mammuthus exilis). Между ними эволюционно располагался мамонт Колумба (Mammuthus columbi), часто ошибочно переводимый как колумбийский. Его зубы содержали 22 — 24 тонких плоских пластины. В ходе эволюции мамонты образовывали региональные популяции, виды и подвиды: ранние, переходные и поздние формы с примитивными или развитыми наборами черт, а также гибриды в местах пересечения ареалов. Это усложнило картину определений, классификаций и датировок, которая изменяется и дополняется так же разнообразно, как и сама их эволюция.
Вхождение в эпоху современности и место гибели мамонтов.
В Евразии обитало стадо мамонтов роду трогонтерии. 800 тысяч Несколько лет назад началось переселение в морозный климат прилегающих к леднику холодных северных степей, богатых высокоэнергетическими злаковыми. В этом приледниковом охлаждении 400 тысячПочти четыредцать тысяч лет назад сформировался последний представитель мамонтов на земле – шерстистый мамонт, или Mammuthus primigenius Blumenbach, 1799.
Зуб его имел самые тонкие и многочисленные пластины. Толщина 24 — 26 пластин часто составляла всего пять миллиметров. Жевательная поверхность образовывала самую мелкую терку среди всех слонов. Это являлось адаптацией к растиранию мелких твердых кормов — летних зерен злаковых и тонких, замерзших до твердости проволоки мелких веточек тундровых кустарников в течение длинной высокоширотной зимы. Шерстистый мамонт заселил огромные пространства гигантской холодной степи, простиравшейся через Северную Евразию и Северную Америку. На этих бескрайних территориях возникали и изменялись популяции, отличавшиеся формами и размерами. Примигениус постепенно уменьшался в размерах после отделения от своего огромного трогонтериевого предка. Более поздние популяции становились все мельче. Пока, наконец, на острове Врангеля на севере Чукотки не оказались самые последние и самые некрупные голоценовые мамонты (то есть послеледниковые; голоцен — текущая геологическая эпоха после оледенения, примерно последние 11700 лет до сегодняшнего дня). Загадка их появления на острове и последующего исчезновения до сих пор не до конца разгадана.

Остров Врангеля стал последним из защищенных мест, где завершались эволюционные пути небольших популяций мамонтов, подобно точкам падения боевых блоков ракеты. Этот остров не был единственным — таковым же стало место на острове Святого Павла в группе островов Прибылова в Беринговом море, где тоже обитали мамонты голоцена, исчезнувшие всего 5600 лет назад. До недавнего времени никто не подозревал о присутствии послеледниковых мамонтов на этих островах и их позднем существовании.

Остров Врангеля расположен в Ледовитом океане, в 140 километрах севернее побережья Чукотки, на меридиане 180 градусов. В 1980-е годы посещение острова было возможно любому гражданину СССР с наличием пограничного разрешения. На остров выполнялись рейсы Аэрофлота. В начале 1990-х годов палеоклиматолог Сергей Левонович Вартанян работал на острове, собрал кости мамонтов и организовал их радиоуглеродную датировку. Открытие ученых стало сенсацией: даты оказались неожиданно молодыми — 8 тысяч лет назад, 6 тысяч лет назад, с самой свежей датой всего 3700 лет назад. Рекордный образец — зуб с полем ОХ-16 и лабораторным индексом Ua-13366, радиоуглеродный возраст 3685 +/- 60 лет. В 1993 году результаты были опубликованы в «Nature» и получили признание мировой науки. Вартанян с коллегами выделили найденных последних мамонтов Земли в отдельный подвид — врангелевских шерстистых мамонтов, mammuthus primigenius vrangeliensis.
В отличие от обычного мнения, подвид примигениуса, названный в честь Врангеля, не является примером островного карликования.
Размер. Не существует четкой границы карликовости, волшебного числа «до/после», подобно условной границе космоса на стокилометровой высоте. Можно считать карликами тех существ, размеры которых вдвое меньше средних или обычных. Высота врангелевских мамонтов составляла 1,8 метра, что отличает их от множества карликовых форм. Например, карликовый мамонт эксилиса (Mammuthus exilis) с высотой в холке 1,2 м, обитавший на островах Ченнэл у побережья Калифорнии (возле будущего американского космодрома, базы Космических сил Ванденберг. При запуске на Марс 5.05.2018 года аппарата InSight ракета-носитель Atlas V проходила на высоте 70 км как раз над этими островами и лежащими там эксилисами).
Карликовые мамонты с острова Средиземного моря действительно были малыми: например, сардинский карликовый мамонт Mammuthus lamarmorae ростом 1,4 м или критский карликовый мамонт Mammuthus creticus, высотой чуть больше метра и считающийся самым маленьким известным мамонтом. По сравнению с этими примерами настоящей островной карликовости, мамонты острова Врангеля были лишь низкорослыми.
Снижение размеров тела в последних популяциях характерно и для материковых мамонтов. Берелехские мамонты в Якутии с возрастом около 12000-14000 лет назад также были низкорослыми, как и синхронные им севские мамонты — последние мамонты Русской равнины, возрастом 13500 лет назад и высотой в холке порядка 2-2,2 метра. Текущий материал не демонстрирует четкого закона уменьшения мамонтов в конце их существования во время завершения плейстоцена. Но в некоторых случаях конечная фрагментация общего макростада на заключительные популяции уменьшала размер мамонтов. Однако до карликовых форм исчезающие остатки материкового мамонтового населения, видимо, просто не успели достичь.
Процесс.Островная карликовость возникает, как и следует из названия, на острове. Однако с врангелевцами было совсем не так. Мамонты на острове жили в разные времена, но карликовости за ними замечено не было. Сначала, в ледниковое время, это были обычные мамонты, не относящиеся к врангелевскому подвиду и жившие 22 — 30 тысяч лет назад, согласно датам, полученным из найденных остатков. Но 12 тысяч лет назад наступила пауза — на следующие три тысячи лет не приходится ни одной находки мамонтов. Значит, эти три тысячи лет мамонтов на острове не было. И только 9 тысяч лет назад появляется сразу плотная череда остатков врангелевцев. Мамонты пришли на остров уже сформировавшейся низкорослой популяцией! Откуда они взялись? Мы пока этого не знаем.
Сергей Вартанян в своей диссертационной монографии «Остров Врангеля в конце четвертичного периода: геология и палеогеография» (С.-Петербург, 2006) пишет, что собрание около тридцати зубов голоценового возраста не позволяет проследить тенденцию к уменьшению размеров мамонтов. Создаётся впечатление, что на остров попала уже сформировавшаяся популяция мелких мамонтов. Неизвестно, где эта группа уменьшилась до заселения острова Врангеля. Возможно, в материковых рефугиумах, которые изолировали её от основного стада, разделенных территориальными фестонами или кое-где вымерших.
Врангелевцы исчезли не постепенно, не угасая в деградации с постепенным уменьшением численности. В последнее тысячелетие до нашей эры (эпоха Среднего Царства Египта или средней бронзы на Дону и Кубани) оставили столько же следов и костного материала, сколько и за предыдущие тысячи лет.
Врангелевцы исчезли не постепенно, оставив в последнем тысячелетии до нашей эры столь же много следов и костного материала, как и ранее. Генетические исследования свидетельствуют о накоплении вредных мутаций из-за изоляции и близкородственного скрещивания при небольшой численности – примерно трехстах особей. В результате неработоспособные гены накапливались в геноме популяции, что привело к ослаблению шерстяного покрова, снижению плодовитости самцов, ухудшению обоняния и, возможно, проблемам с пищеварением.
Несмотря на генетические проблемы, популяция мамонтов оставалась стабильной по численности. Об этом свидетельствует частота дат и большое количество остатков последнего периода существования вида. Мамонты вновь проявили свою удивительную эволюционную пластичность. Какое катастрофическое краткосрочное событие или череда событий привело к быстрому исчезновению последних мамонтов Земли, остается загадкой. Возможно, это были две-три кризисные зимы, крайне сложные для добычи корма. Или эпидемия, поразившая последних представителей вида. В отличие от большинства континентальных популяций, врангелевский подвид, скорее всего, не подвергался давлению охоты со стороны человека. Следы проникновения на остров древних людей обнаруживаются лишь спустя 800 лет после исчезновения последнего мамонта. Таким образом, около 3700 лет назад на острове Врангеля завершилась долгая история мамонтов на Земле, оставив множество своих тайн нераскрытыми.
История продолжается.
История последних мамонтов не завершилась, а продолжается неожиданными и удивительными результатами, полученными в 2021 году. Эти результаты кардинально меняют наше понимание исчезновения шерстистого мамонта. Это лишь начало потока информации, который будет расти, приводя к новым открытиям.
Новые методы выявления останков мамонтов в разных местах меняют наше представление о том, как вымерли мамонты.
Макроостатки мамонтов — фрагменты тел: кости, зубы, бивни, мягкие ткани и так далее — обнаруживают визуально на местности или в грунте. Принадлежность остатков мамонту определяют по форме и виду, например, характерный отщеп у бивня не свойственен мышам или песцам. Из полевого образца отпиливается кусок для радиоуглеродного метода — лабораторный образец. Из материала образца делают фенолы, которые анализируют на количество атомов радиоактивного углерода. По количеству атомов определяют возраст образца.
Современные методы секвенирования ДНК позволяют находить следы мамонтов и других животных иначе. Вместо анализа отдельных костей или останков берётся образец древней почвы.
В нем содержатся фрагменты ДНК различных животных и растений, живших во время формирования этой палеопочвы. Затем проводится массовое секвенирование всех ДНК из образца. Многие ДНК восстанавливаются достаточно полно для достоверного определения семейства или рода носителя. В одном образце палеопочвы идентифицируются ДНК слонов, оленей, зайцев, мышей, носорогов, волков и других животных.
Для определения возраста останков нужно отправить органику из палеопочвы на радиоуглеродную датировку. Полученный возраст почвы покажет время жизни животных, оставивших ДНК. Это упростит определение вида мамонта: если возраст образца 10 или 15 тысяч лет назад, а ДНК указывает на семейство слонов, то это шерстистый примигениус, обнаруженный в месте формирования палеопочвы. По самому молодому образцу с ДНК мамонта можно определить дату последнего его появления в этом месте — LAD (last appearance date).
Группа исследователей под руководством известного датского ученого Эске Виллерслева провел масштабное и трудоемкое исследование. Eske WillerslevСобрано 535 образцов палеопочв с 74 участков приполярных районов Евразии и Аляски для поиска дат последнего появления различных животных. Результат по северу Таймыра оказался неожиданным. опубликованДвадцать октября прошлого года в журнале «Nature» появилась статья о том, что последние мамонты на Таймыре жили всего 3900 +/- 200 лет назад. Это значит, что континентальные мамонты обитали на полуострове до середины голоцена! Разница их исчезновения со временем исчезновения популяции врангелевских мамонтов составляет лишь 200 лет — ровно на величину ошибки +\- 200 лет.
Жизнь и вымирание мамонтов на Таймыре и острове Врангеля происходили одновременно. Врангелевская популяция является сейчас последней на Земле лишь формально, разделяя с таймырской всего 200 лет — период, легко превосходящий погрешность датировок. Это ставит под сомнение статус врангелевской популяции как последней на Земле.
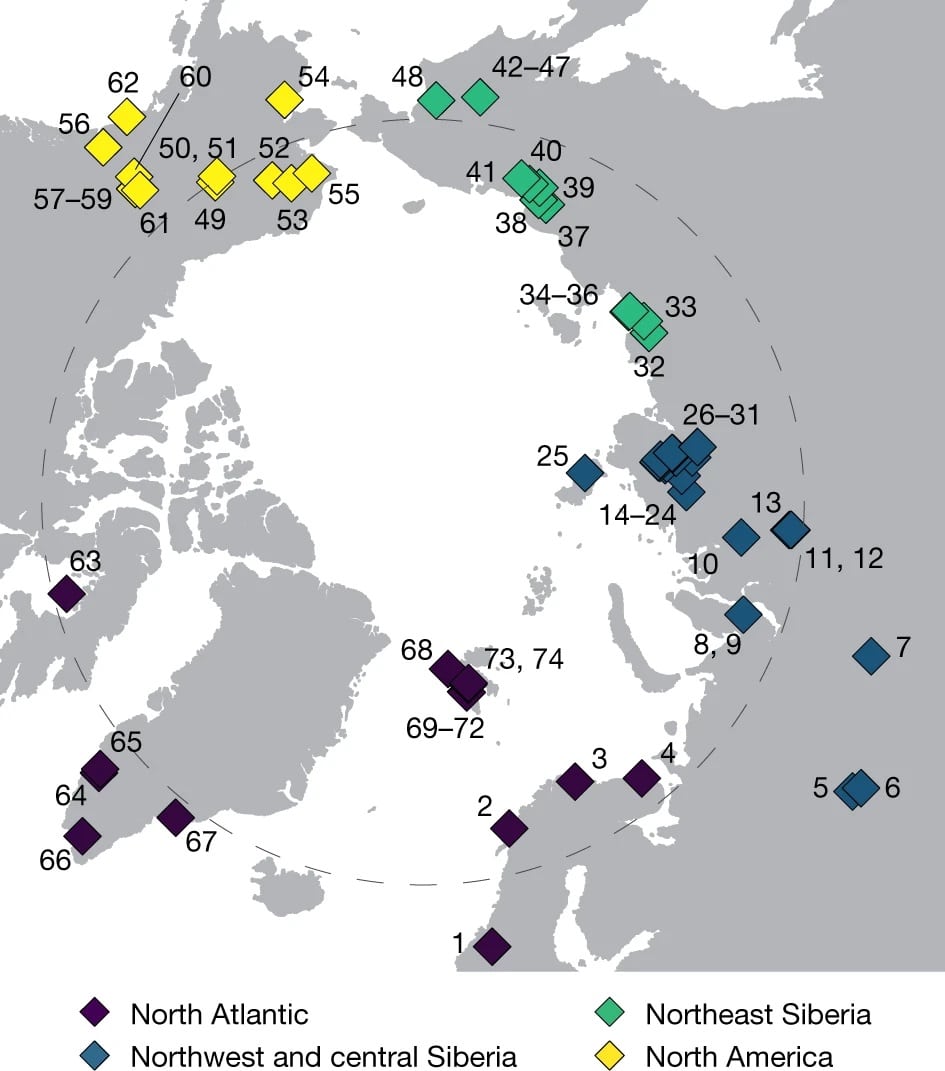
В декабре 2021 года, через полтора месяца после публикации первой статьи, в журнале «Nature» появилась другая. статьяРезультаты аналогичных исследований в области центрального Юкона, проведённые другой группой исследователей, показали достоверный ДНК-сигнал присутствия мамонтов в образцах возрастом 5700 лет. Эта дата древнее последних врангелевских мамонтов всего на две тысячи лет. Таким образом, и на канадском Юконе обнаружены континентальные североамериканские мамонты из середины голоцена. Такой временной промежуток жизни мамонтов никто не мог предположить по традиционным макроостаткам.
Новые исследования предстоят. Площади ареалов мамонтов на материках значительно превышают ареал мамонтов на острове Врангеля. Животное за свою жизнь оставляет неизмеримо больше своих ДНК-следов на местности, чем собственных останков. Ведь оно проходило сотни и тысячи километров, а ДНК в виде отмерших клеток ежедневно и ежечасно оставалось на пройденном пути. В помете, слюне, шерсти, стираемых об землю частицах бивня, кожи стоп и других микроостатках и выделениях. Увеличение территориальной плотности отбора образцов палеопочв, количества образцов и быстрое совершенствование методов массового секвенирования ДНК из почв открывают неведомые ранее перспективы.
Наблюдения сигналов голоценовых мамонтов будут расширяться – подобные исследования только начинаются. Выводы о разных континентальных популяциях голоценовых мамонтов показывают, что климатические условия и ландшафтная кормовая база для них сохранялись широко до середины голоцена, всего лишь несколько тысяч лет назад. Поэтому ожидается омоложение последних популяций мамонтов на обоих континентах. Становится ясно, что с последними из мамонтовых могикан пока всё не выяснено. Впереди нас ждут дальнейшие, и скорее всего удивительные открытия.